 At first glance, these topics seem like child's play. Breeding brings new varieties, so it is per se a contribution to diversity, which everyone loves anyway. All's well that ends well; everyone's happy. On closer inspection, however, the situation is quite different: breeding means selection and reduction, so strictly speaking it is pretty much the opposite of diversity. This is reinforced by the mechanisms of agro-industrial production and the market (more of the same is better). And finally, the new breeding technologies fit into the picture, producing more variants of the same. And if you look around, you'll find that people loathe diversity pretty much to the same extent that they praise the concept. And this is somehow both strange and exciting.
At first glance, these topics seem like child's play. Breeding brings new varieties, so it is per se a contribution to diversity, which everyone loves anyway. All's well that ends well; everyone's happy. On closer inspection, however, the situation is quite different: breeding means selection and reduction, so strictly speaking it is pretty much the opposite of diversity. This is reinforced by the mechanisms of agro-industrial production and the market (more of the same is better). And finally, the new breeding technologies fit into the picture, producing more variants of the same. And if you look around, you'll find that people loathe diversity pretty much to the same extent that they praise the concept. And this is somehow both strange and exciting.
Diversity is what everyone is talking about. After the term and concept of diversity have played an important role in biology since Darwin, the demand for diversity is currently being transferred to many other areas of knowledge and life, where diversity is not infrequently elevated to a new bogeyman that alone defines what is politically correct. Those who do not live diversity to the very last crevices of their lives and, above all, do not push it as far as grammar, already seem to violate the new laws. The fact that this exclusion (of what is not politically correct) could impair, even deny the imperative of diversity itself occurs to very few people. But it is precisely this contradiction that diversity is difficult to think through, at least for people, and that it sometimes turns into the opposite, is probably characteristic of diversity – and also shapes the following considerations.
Our topic: diversity and breeding
Of course, we don't want to get lost in generalities here but rather report specifically on our topic: diversity and breeding. To what extent is it possible to allow as much diversity as possible in breeding? If diversity is a basic principle of life that makes life possible in a sustainable way, and if breeding crops is a task that is essential for survival in a future with 10 billion people, then we must allow as much diversity as possible in breeding. Then I think we need to bring breeding and diversity together. Successful breeding needs as much diversity as possible, and should also allow as much crop diversity as possible.
Why diversity in plants and crops?
Why exactly should diversity be good and productive in nature, especially in plants and crops, but actually also in humans and animals? The answer is simple: because diversity maximises the chances for survival. The more lots you have, the greater your chance of winning the big prize. The more forms of life that are formed in plants and animals, the greater the chance that there will be adaptable and survivable units among them that will succeed and survive, that can adapt to the prevailing circumstances.

Diversity is certainly the dominant strategy. But I would also like to point out here that there are also concepts of sameness that don't seem to work so badly: military organisations are certainly not crazy about diversity; expats meet with their peers to strengthen their identity; nationalism and xenophobia are quite successful, but also less sympathetic concepts, at least on a short time axis. And how many of us have wished for clones – preferably of ourselves, of course. In plants, too, there are obviously synergistic mechanisms and information exchange between the same or similar plants that promote life in association, as equals among equals. Perhaps here, too – in these counter-concepts only briefly touched upon – the dialectic that somehow seems to be inherent in diversity becomes apparent. Diversity is clearly positive, it helps life to survive, but underneath there are many counter-concepts that at least partially work as well – or even diametrically oppose diversity.
Human behaviour in relation to plants is anti-diverse
Do you want to hear an example? Quite simply, our human or more generally animal behaviour towards plants is fundamentally anti-diverse. The proverbial bear that roamed through the fruit forests of Kazakhstan in primeval times and always ate only the largest and sweetest apples, thus preforming today's cultivated apple – this bear had a plan: he wanted to use the apple to his advantage, and so he chose the largest and sweetest. Animals and humans select; they select according to certain preferences, which do not necessarily have to be in the interest of the plant; they select, they sow, they transport, they clone at some point, they reduce diversity to a very large extent with their conscious and unconscious actions. This animal behaviour is only the preliminary stage of the current Anthropocene, where man seems to be completely abandoned by his natural senses, and plunders and destroys what nature still gives.

As always with the human being, who is a rather clever animal, this all often works a little more complicated and perhaps also better camouflaged – namely as self-deception. Water is preached – wine is drunk. Few concepts are currently more popular in politics and administration than singing the praises of diversity. Old varieties are collected for all they're worth and until the fields burst – but at the same time, officially guarded variety lists limit access to cultivation. And when – essentially man-driven – our framework conditions, climate and environment change, when old familiar plants leave and new ones immigrate, then the hunt for invasive neophytes is quickly sounded, as if there were nothing more important than to suppress any help for a serious diversity strategy in the bud. Not infrequently, in the same paragraph, indeed in the same sentence, the absurdity is taken to extremes and one can then read that neophytes would almost naturally destroy diversity, which is quite simply a popular lie. Not only is everything new that comes along basically an enrichment of diversity, but countless island studies show that immigrating animals and plants do not threaten biodiversity, but enrich it. But perhaps it is necessary to make it clear once again and clearly at the end of this section, just so that no misunderstandings arise: plant bans, which neophyte policies always amount to in one form or another, harm diversity and do not benefit it in any way. If you ban plants, you ban diversity. That’s a fact.
Breeding is anti-diverse
And now we are getting pretty close to our actual topic: of course, breeding is also anti-diverse. It is selection and propagation. It may increase diversity at first (in crossing, in populations, in sifting genetic diversity), but after that diversity goes rapidly downhill. The breeder who does not reduce his/her young plants will suffocate in the jungle of plants and will get lost in the infinite breeding fields. I remember a visit to an apple breeder in the Czech Republic when we stumbled through a 30-year-old breeding area that was full of molehills and foxholes; I permanently wondered why the breeder did not finally dispose of this material and reduce it. Breeding increases diversity – in order to then reduce it again as quickly and efficiently as possible. This shows once again the dialectical character of diversity, which we have already encountered in various places in the previous train of thought.

Picture: Which healthy plant would you choose?
How is more diversity in breeding possible?
Once again, if diversity is a central survival element of life, and if breeding in cultivated plants aims to do precisely the same thing, namely to promote the survival of plants and humans, then it is probably logical to ensure as much diversity as possible in breeding. In the following, based on our experience at Lubera, I will show 10 strategies on how to increase diversity in breeding – always knowing that in the end, breeding also has a strong anti-diversity flavour. If breeding neglects the diversity side, it not only loses its appeal as an art, it also loses its creativity and also the chance to create something truly new.
1. Multicrop breeding programmes instead of specialisation
I am convinced that multicrop breeding programmes are superior to greater specialisation. What you lose in depth, you gain in breadth and stimulation. In our 20-30 large and smaller breeding projects, I always notice that the projects cross-fertilise, that slowly but surely an approach emerges that works quite well even with very different plant species. I first noticed this externally, so to speak, when I visited the Terra Nova perennial breeding company south of Portland, Oregon in the United States: the entire ornamental plant world puzzled and puzzles over how the colleagues manage to bring 15-30 new perennial varieties to market every year. Quite simply, this is achieved thanks to a distinctive multicrop strategy. Each breeder in the breeding team works on several different species, experiences and approaches are shared and duplicated if possible. Another hint that my statement – multicrop is better than specialisation – might be true is an experience that many breeders share or at least like to report about other breeders: above a certain size, the effort/yield ratio in breeding tips (more young plants equals more results equals more varieties) and develops into an inversely proportional breeding nightmare: the more breeding is done, the larger a breeding programme becomes, the more it approaches the point where the possible yield continuously decreases. The more diversity I produce, the more young plants I have in my fields, the harder it becomes to select them back down. But it is not only the law of large numbers that can no longer be mastered (or only statistically or only with markers); in my opinion, it is really also the sheer lack of ideas that can spread in very large and quantity-fixated breeding programmes. The breeder becomes the image of his/her monoculture. Well, I readily admit that I exaggerate a bit here…but just a bit. The most exciting breeders I have met have always worked on several species! And two or three as a hobby as well.
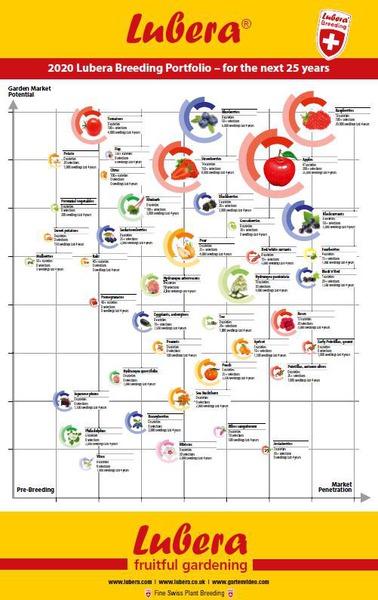
Picture: Lubera breeding portfolio
2. Combination breeding with free recombination of genes vs. mutation breeding and other variant breeding methods
The apologists of newer breeding methods such as CRISPR/Cas often argue that mutation breeding has been done before, also with artificial methods (colchicine, radiation), and that it is therefore difficult for them to understand why these new “genetic engineering” methods are not allowed now. We do not want to have this discussion here, but I would like to turn the argument around: all, or at least most, of the new breeding methods such as CRISPR/Cas are mutation breeding methods; they only produce variants of the same thing. Yes, of course, colleagues will now counter, that may be so, but if we can make the valuable variety x even more valuable by adding resistance trait y, then that is only to be welcomed, isn't it? Yes, but what was that again about diversity? All these methods definitely do not lead to more diversity, but to less (or let's say micro-diversity of the collected mutations). Successful varieties become even more successful, are cultivated even more, the monocultures become bigger – this goes on just as long until the mounted resistance is broken...
Only classical combination breeding, which takes plant sex as a basis, lets genes combine freely and really produces new plants or diversity. I'm really afraid that this basic truth will be forgotten because of the greed for methods in agronomy.

Picture: classic crossing; the female organs of the flowers are pollinated with male pollen
3. Against the temptation of hybrid breeding
Of course, I am not quite as bold as the title of this section promises. Of course, I too must acknowledge that hybrid breeding contributed significantly to the advances in agriculture and food production in the 20th century (the first green revolution). The basic concept of hybrid breeding is this: inbred lines fix traits. By crossing inbred lines from the 8th to 10th generation, these traits can be ideally combined and the heterosis effect (the strangeness, so to speak, of distant inbred lines) leads to an explosion of many mainly quantitative traits (fruit size, fertility, yield, etc.). Actually, one cannot say very much against it, the results speak for themselves.
Only: of course, the hybrid breeding method works diametrically against the goal of diversity. Everything is aimed at crossing out diversity, standardising traits, reducing the genetic range, and then in a final step, during the production of the so-called F1 generation, to partially broaden it again in a controlled and planned manner (but please not too far...).
You have surely noticed, as a trained engineer I must not dare to rail against hybrid breeding, but I would like to note at least cautiously that the reversal of hybrid breeding can also enable creative breeding in many species. In tomatoes, for example, we try to break hybrid breeding by splitting hybrid varieties, then crossing valuable individuals, doing single plant selection, and vegetatively propagating the interesting variety candidates. In this way, I do not need my limited resources for maintaining the costly inbred lines, but can always cross new diversity in each breeding generation and stabilise it via vegetative propagation (if desired). As you can see, even successful and at first sight, unbeatable breeding concepts can easily be turned around, often to the benefit of diversity.

Picture: Tomato breeding field at Lubera in Buchs (CH)
4. Old varieties as a source of diversity? Rather not!
The last 30 years have been the age of old varieties in many crops and many regions of the world. Old is good, and always worth promoting. This is probably because in the end you already know it and don't have to expect unpleasant surprises. In Switzerland, for example, gooseberries were collected, which, with two or three exceptions, are all no longer cultivable because of the strong susceptibility to powdery mildew. The example also shows one of the main problems of the old varieties well: they are usually old and forgotten because they have simply been replaced by better varieties.
Interestingly, there are also hardly any successful breeding projects that are really based on old varieties, which have explicitly made it their task to help the “old” back to its ancestral right. Most of the time, such attempts to cross old varieties and thus to fertilise the new are quite a failure: I have observed this several times with apple breeding colleagues, where after a few years the entire “old” crossings and crossing results have disappeared into oblivion. Somehow this is also logical: the modern current varieties carry the genetic material of the old varieties (which are mostly only 50-200 years older) on their backs, so in most cases, nothing really new can develop…
The only exception I can think of is the rose breeder David Austin, who explicitly wanted to combine the charm and fragrance of the old varieties with the continuous flowering ability and the colour spectrum of the modern varieties. I have been to the breeding fields of David Austin several times – and I suspect, probably rightly, that perhaps in the early years the old was really combined with the new, but that in the last 50 years modern and more and more modern varieties were bred, into which new resistance properties were also deliberately bred. This is because old varieties very often fail in resistance, either because disease pressure increases in monoculture or because our demands on the product (without spots or for fresh enjoyment rather than processing) have changed.
Nevertheless, time and the historical amount of plant individuals produce mutations that then “overwinter”, so to speak, in old varieties. As uninteresting as it is to expect a boost for modern breeding from breeding in old varieties, it can always be exciting to pick up individual traits conserved in old varieties and incorporate them into modern breeding. In the case of apples, we started breeding old American cultivars of the 19th and 18th centuries that produce seedless apples over 10 years ago. The underlying mechanism here consists of two parts: the seedless apple varieties do not produce petals, but they have reinforced sepals. The lack of petals means that the flowers, which are thus rather unattractive, are not approached by any or almost no insects. This would be the first mechanism. In addition, these varieties are fully or even partially able to set and develop fruit parthenocarpically, i.e. without fertilisation. Presumably, this is again related to the strengthening of the sepals and the receptacle.

Bild: seedless apple 'Faibella' – the seedless apple without seeds or core, a botanical curiosity - or the future of the apple?
This somewhat abstruse diversity would of course not last very long in nature (at least not as a dominant trait), as the ability to reproduce would be quite limited. This is yet another irony that seems to accompany us at every turn when it comes to diversity...

Picture: Flower buds of the seedless varieties, with reinforced sepals but without petals
5. Species hybrids
Species hybrids are – in breeding and also in nature – a quite successful way to bring more diversity into play, to enlarge the field of possibilities, so to speak. For me, the most beautiful example are the different citrus species and types. From only four or five original species, the whole variety of forms that characterises the citrus world seems to have unfolded: let us think, for example, of the finger limes, then of the miniature kumquat, only 1 cm in size, and on the other side the giant grapefruits or grapefruit hybrids. This is diversity pure. One condition of the possibility of this diversity is surely that a genus is always richly endowed with promiscuous abilities so that there are few barriers to hybridisation. This is particularly the case with citrus; even related genera such as Microcitrus or Fortunella and Poncirus can be hybridised with citrus or hybridise themselves in a natural encounter.
Nevertheless, when you try to create something new, in the free middle, so to speak, with species hybrids, it usually fails. The jostaberry is not really a great success (although we are also breeding there), and also tayberries as well as other Rubus hybrids always suffer a bit from the fact that they are neither fish nor bird. In the case of Rubus, blackberries and raspberries, there is also the fact that the hybrids have a particularly difficult time with the basic problem of all Rubus species: does this strange cone in the middle, which stabilises the aggregate fruit, really belong to the fruit or not?
More promising than the production of a new fruit, a new hybrid species, so to speak, seems to me the goal of bringing individual special characteristics from one species into the other. For example, we are trying to bring the autumn bearing ability of autumn raspberries, which has been extremely developed in the last 50 years, into blackberries one more time (this happened accidentally/naturally once before at the beginning of the 20th century).
Transfers could also be made between gooseberries and blackcurrants, enriching both sides: resistance to powdery mildew is now much better established and more widely secured in blackcurrants than in gooseberries. And especially the largest gooseberries could still contribute to the size development of the cassis fruits.

6. Back to the origin!
Since Vavilov (Nikolay Ivanovič Vavilov) it belongs to the basic task of a breeder to go back to the origins, to the wild diversity or even to the semi-domesticated diversity as basically as possible or even in between. Vavilov had conceived the concept of genetic centres early in the 20th century, recognising that the origins of a species are usually found where the greatest diversity is found. Yet the breeding reality is quite ambivalent: one wants to and should go back to the original diversity, but one doesn't really have the time. As a result, in many large breeding programmes, pre-breeding, as it is often symptomatically called, is outsourced to a special organisation and agency. I think this tends to be a mistake: if you see pre-breeding as just integrating some special features (e.g. a new resistance), you are likely to miss more opportunities than you take. The classic picture that breeders like to paint, after all, tells the story that the wild varieties first have to have their accumulated negative traits, their fluff, exorcised during domestication. But probably, and more productively, the glass should be seen as half full (not just half-empty) when domesticating wild species: in addition to the countless negative traits (fruit size, uneven ripening, self-fertility, etc.), there are always new and exciting traits to be discovered, which a modern market may be waiting for. For example, we have crossed a hanging wild apple (a Malus species hybrid) with apples about 10-20 mm in size into our breeding programme – and already after two generations of breeding we see a real wealth of new interesting apple traits:
• Yellow flesh, but also all shades from red to orange
• New flavours, super fruity
• Scab resistance
• Resistance to Marsonina
• All plant structures from hanging to normal
We are almost sure that already in this generation we will be able to select new, exciting varieties for the home garden, maybe already for the niche market.

Another important variation of going back to the origin is the development and domestication of entirely new species, or of semi-domesticated species. This area is systematically underdeveloped in the current breeding landscape because it does not promote what does not yet exist on the crop map. This, too, is ultimately based on an anti-diverse short-circuit: both within companies and on the part of governmental funding institutions, what does not yet exist has no chance. This is itself an important factor in the drying up of crop diversity. Crops are regularly dropped because they no longer fit climatically or because consumption habits change decisively; but almost no new crops are added either, at least in temperate climates...
In the following table, we present some of the Lubera® domestication projects we are currently working on. We have just described the crucial problem above: it is difficult to assess what does not yet exist and therefore one tends to be conservative when making investments. With semi-domesticated species such as Lonicera and Amelanchier, there is at least one tendency.
| Species | Wild | Semi-domesticated | Target characteristics for domestication |
| Ribes aureum, Fourberries |
x | Self-fertility, fruit size, uniform maturity, crop stability, wood stability vs. perennial character | |
| Lonicera caerulea, Haskaps, Firstberries, Mayberries | x | Fruit size, sugar/aroma, match early maturity with healthy foliage of later genetics, self-fertility | |
| Oca, Oxalis tuberosa | x | Adaptation to Central European conditions, slightly more heat tolerance, long day tuber formation | |
| Amelanchier alnifolia, Saskatoonberries | x | Fruit size, fruitiness, uniform ripening, number of berries per cluster, harvestability, no sap on stem |
A good indication of future rising stars could be the success story of the cultivated blueberry, which has completed its domestication career within a good 100 years from wild picking in the American eastern states to becoming the world's second-largest berry species. Why was and is the blueberry so successful – and why is it on the verge of also replacing the strawberry as the queen of berries?
- Blueberries are everybody's darling, everybody likes them, but they are also not incredibly characteristic, they don't cause any problems
- They genetically offer and offered the basis for the cultivation in almost all climatic zones, thus for the year-round supply of the consumers and the trade
- They are easy to harvest and transport
- They are “clean”, uncomplicated, the fruit is formed like a pill, but healthy 😉
7. The dead angle assistant for growers

Growers know their own breeding material best – and know how to use it best. The effect of this process is often underestimated (I underestimated it too), but it gains strength year after year, decade after decade. After 20-30 years, breeders have their breeding lines, successful parents and other tricks together – and they are already in danger of becoming business-blind. Here, too, only diversity can help. For example, in the second stage of apple breeding we have a constantly updated apple assortment. Our main focus is not on current assortment varieties, but on as many current novelties from as many geographical regions as possible. This not only helps us to develop our own genetic material, but also sharpens our judgment: we are in the top group for this trait, and we need to catch up for that one. The rule that 50%, but at least 30%, of crosses should be made with foreign genetic material belongs in the same chapter.
8. Against the logic of agricultural markets – the bottom-up approach

Picture: Blue = "diverse" potato variety with high wild content and new traits, Purple = further developed niche varieties with improved agronomic traits, Red = varieties for the wholesale market.
Bottom-up approach: Lubera first breeds varieties for the "diverse" home garden market. For apples, raspberries, blackberries and potatoes, niche varieties will then be developed.
The industrial agricultural markets function according to a purely industrial logic: more of the same improves efficiency and increases profit margins, especially in the area downstream of actual agricultural production. This applies to the major agricultural crops such as potatoes, soybeans, corn and cereals, but also to vegetables and fruit. The buyer, who made two successful actions with Gala apples last year, will try in his/her logic to make three actions this year. He/she then wants to use this successful variety also in the organic market, although it is not at all suitable for organic production and needs 20+ sprays for good fruit quality. Finally, he/she will also start with promotions in the organic market and so on. (To clarify: Gala is really by far the largest variety in organic production in Switzerland, for example...). This system, of course, causes anything but diversity; it works diametrically against it.
We, on the other hand, have developed a bottom-up approach with our breeding at Lubera: we breed first for the home garden market, for which novelty and also diversity and otherness are a value in and of themselves. At the same time, one must not be deceived: for many fruit and berry species, but also for vegetables, these amateur markets are large to very large, in some cases much larger than the plant market for agricultural cultivation. Lubera is only trying to develop varieties for the agricultural niche or large market in the medium term for a few species, apple, pear, raspberry, blackberry and potato. We also hope to bring more diversity into the agricultural markets through the bottom-up approach (home garden market first, niche market second).
When we submitted a research project on potato breeding to the federal government (in Switzerland) together with an agricultural training centre a few months ago, the rejection came soon after. Here was the reason: we would not have been able to show how the results of our project (that involved cross-breeding Phytophthora-resistant wild potatoes and semi-domesticated potatoes) could meet the quality criteria of Swisspatat (the potato industry organisation). This refusal is, in fact, the exact explanation of why the potato always looks like a potato and why, to this day, 150 years after the Irish and European potato famine (after the blight invasion of Europe), there are still no truly resistant varieties on the market.
9. What does marker-assisted breeding have to do with Easter egg hunts?
Two years ago, at a blueberry congress in Trento, Italy, I listened to an Italian breeder colleague: she had tried to define what exactly is good fruit quality in blueberries (firmness, size, sugar content, acidity, etc.) and for these traits, she was looking for so-called molecular genetic markers to be able to select for quality at an early stage, already in a small seedling, two to three years before the plant would be able to bear fruit. Of course, this approach is fascinating for all breeders, who already from the age of 40 include their current age in the calculation of the still possible breeding cycles: for every 7-10 years of a breeder's life, 1 crossing cycle, one generation is possible in the case of apples, in the case of blueberries maybe every five years. And the marker method (I define a desired trait and then look for a genetic marker for it) can and will work, earlier for simple traits, slower for more complicated ones. The only problem with this is that you will only find the Easter eggs that you have hidden before. You will definitely never be able to find a completely different definition of fruit quality than the one my Italian colleague defined for blueberries. In other words: you will always find the same thing...

Picture: Breeder hare finds the eggs he hid himself
By the way, this does not speak against the use of marker-assisted breeding in breeding, we also use markers for tomatoes, apples, potatoes and maybe soon for raspberries and blackberries. But the epistemological Easter egg problem also clearly shows that one should not rely on markers too early in the selection process.
10. Phenotype before genotype

And this would then be the solution to the epistemological problem formulated above under point 9: phenotype before genotype. Before we turn to the genetics of a plant, the breeder must always look at how it behaves in the field, and of course in the palate in the case of edible plants. Now this sounds like a no-brainer, but it is not; the speed trial of markers is far too great for that for breeders, especially academic breeders. When we started breeding tomatoes a few years ago, we also thought that we wanted to screen our breeding population quickly and also systematically with markers for the known resistance species Ph1, 2 and 3. Punch small leaf pieces out of the leaves, send them in and I know which varieties are resistant and which are not. Fortunately, the markers and the tests did not work so well...Anyway, we had enough time to observe all the plants in the field. In fact, it turned out that another breeding line completely without the already known resistance genes Ph1 to 3 was much more interesting and also much more resistant in the field than the marketed genes. We would never have gained these insights without the close examination of the phenotype in the field. There simply aren't as many shortcuts in breeding as breeders would sometimes like. Instead, there are distractions and surprises galore. In my opinion, it is not a very good idea to exclude chance, the driver of diversity, from breeding.
This contribution was given by Markus Kobelt as a lecture at the Rheinhof in Sennwald, Switzerland (Regional Agricultural Training Centre).
